VgrS受體細(xì)菌組氨酸激酶在NYG培養(yǎng)基中的的生長(zhǎng)曲線圖
革蘭氏陰性細(xì)菌作為一種單細(xì)胞生物,這類細(xì)菌在感知外界刺激的過(guò)程中,主要利用細(xì)胞內(nèi)膜上的受體監(jiān)測(cè)環(huán)境信號(hào)。其中,受體組氨酸激酶以蛋白可逆磷酸化方式(磷酸化-脫磷酸化)完成環(huán)境信號(hào)的跨膜傳遞和信號(hào)轉(zhuǎn)導(dǎo),發(fā)揮著類似高等動(dòng)物中樞神經(jīng)系統(tǒng)的作用,因而被研究人員形象地稱為細(xì)菌的“智商(IQ)”。本論文的研究人員發(fā)現(xiàn)細(xì)菌周質(zhì)空間蛋白酶以不可逆、高度特異的方式修飾細(xì)菌的“IQ”,促進(jìn)細(xì)菌抗逆脅迫水平。該研究從遺傳學(xué)、酶學(xué)和生物物理學(xué)方面提出了充分的證據(jù),證明上述過(guò)程對(duì)于細(xì)菌適應(yīng)脅迫環(huán)境的重要意義。
實(shí)驗(yàn)步驟
使用Biosense微生物生長(zhǎng)動(dòng)態(tài)監(jiān)測(cè)系統(tǒng)測(cè)試了不同類型的細(xì)菌及細(xì)菌突變體在NYG培養(yǎng)基中的的生長(zhǎng)曲線圖,培養(yǎng)維溫度為37度,培養(yǎng)時(shí)間為18-84h,每隔30分鐘測(cè)試一次OD600值,培養(yǎng)過(guò)程中搖晃、生長(zhǎng)及OD值的測(cè)試全是自動(dòng)完成的,從而能夠長(zhǎng)時(shí)間準(zhǔn)確的測(cè)試出不同類型的細(xì)菌在不同條件下的生長(zhǎng)情況,獲得相應(yīng)的生長(zhǎng)曲線圖。
實(shí)驗(yàn)結(jié)果
在細(xì)菌細(xì)胞中,組氨酸激酶(HKs)是一種受體,可以監(jiān)測(cè)環(huán)境和細(xì)胞內(nèi)刺激。HKs及其同源反應(yīng)調(diào)節(jié)因子構(gòu)成兩個(gè)部分信號(hào)系統(tǒng)(TCSs),通過(guò)可逆的蛋白質(zhì)調(diào)節(jié)細(xì)胞的穩(wěn)態(tài)磷酸化。然而在病原細(xì)菌-野油菜黃單胞菌感染植物的過(guò)程中,有一個(gè)名為VgrS的受體組氨酸激酶發(fā)揮著關(guān)鍵作用。VgrS感知寄主體內(nèi)由固有免疫反應(yīng)產(chǎn)生的缺鐵環(huán)境,調(diào)節(jié)病原細(xì)菌從寄主中爭(zhēng)搶和吸收鐵的過(guò)程。VgrS突變或失活后會(huì)導(dǎo)致細(xì)菌鐵代謝紊亂,致病力嚴(yán)重下降。然而,VgrS的功能具有兩面性。當(dāng)細(xì)菌面對(duì)高滲脅迫時(shí),VgrS信號(hào)通路的激活會(huì)嚴(yán)重阻礙細(xì)菌的生長(zhǎng)。為了迅速淬滅該通路,細(xì)菌周質(zhì)空間內(nèi)的蛋白酶Prc結(jié)合到VgrS位于氨基端的信號(hào)感知區(qū)域,隨即以一種“掐頭不去尾”的方式,特異性切割前9位氨基酸并且抑制VgrS的激酶活性。VgrS被Prc特異性切割失活是細(xì)菌對(duì)抗高滲脅迫的前提條件。
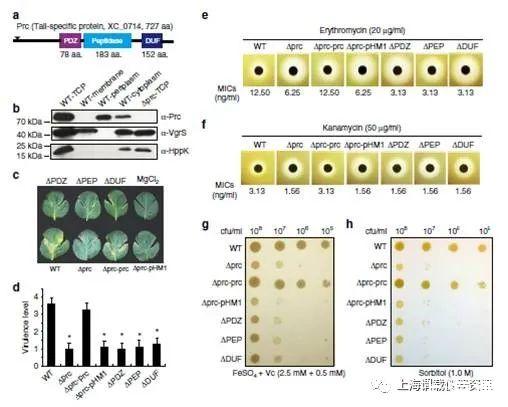
圖1、圖a表示的是Prc蛋白質(zhì)二級(jí)結(jié)構(gòu)示意圖。圖b表的是Prc蛋白是位于細(xì)菌周質(zhì)和細(xì)胞溶膠中,采用的是采用Western blot技術(shù)檢測(cè)不同細(xì)胞組分的Prc蛋白。圖c表示的是Prc的失活導(dǎo)致了細(xì)菌的毒力的不足。圖d表示的是在圖c中對(duì)應(yīng)的菌株的毒性評(píng)價(jià)圖,采用了半定量標(biāo)準(zhǔn)測(cè)定了菌株的毒力水平。圖e、f表示的是該突變體對(duì)多種抗生素(紅霉素e和卡那霉素)敏感性。圖g表示的是Pcr的失活導(dǎo)致了對(duì)鐵脅迫的過(guò)敏,菌株的培養(yǎng)是在含有2.5mM FeSO4+0.5mM維生素C的NYG瓊脂上,于28℃條件下培養(yǎng)72 h。圖h表示的是Prc的失活導(dǎo)致了對(duì)滲透應(yīng)力的過(guò)敏。
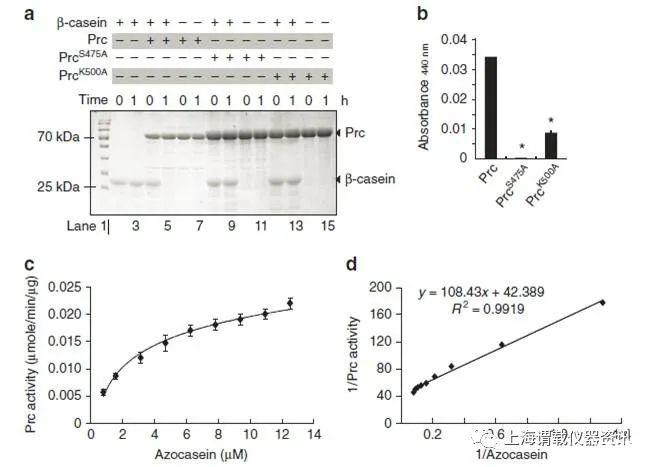
圖2、Prc是一種絲氨酸內(nèi)肽酶。Prc可降解β-酪蛋白,其內(nèi)肽酶活性依賴于Ser475和Lys500。圖b表示的是通過(guò)降解底物的偶氮雜蛋白酶定量研究Prc內(nèi)肽酶活性。圖c和圖d表示的是偶氮酪蛋白水解酶活性的邁克爾-門登動(dòng)力學(xué)擬合曲線。

圖3、Prc通過(guò)切割VgrS的敏感區(qū)域來(lái)抑制其自身激酶活性。圖a-圖c表示的是Prc以蛋白酶活性依賴的方式抑制全長(zhǎng)VgrS的自磷酸化過(guò)程。圖d-圖e表示的是Prc在體外降解了VgrS感知寄主體。圖f表示的是Western blotting顯示Prc在體內(nèi)降解n-末端HA-tags。

圖4、vgrS n端缺失抑制了突變體在滲透脅迫下的生長(zhǎng)缺陷。圖a、c、e和g表示的是細(xì)菌在NYG培養(yǎng)基中的生長(zhǎng)曲線圖。圖b、d、f和圖h表示的是細(xì)菌在NYG培養(yǎng)基加上1.0M山梨醇的生長(zhǎng)曲線圖。用丹麥Biosense微生物生長(zhǎng)動(dòng)態(tài)監(jiān)測(cè)系統(tǒng)測(cè)定其其相關(guān)的生長(zhǎng)曲線分析圖。每個(gè)數(shù)據(jù)點(diǎn)是六個(gè)樣本的平均值。
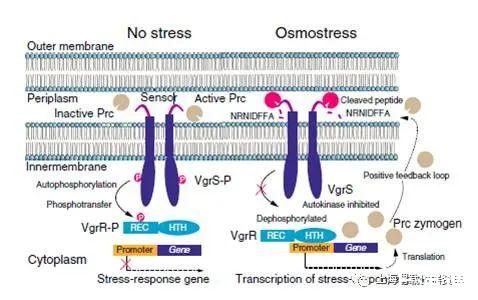
圖5、VgrS蛋白水解引起的細(xì)菌應(yīng)激反應(yīng)模型。當(dāng)細(xì)菌在滲透脅迫下生長(zhǎng)時(shí),Prc被激活并分裂n端肽(NRNIDFFA)對(duì)VgrS的感測(cè)區(qū)具有抑制作用,導(dǎo)致后者的自身磷酸化。
總結(jié)
研究發(fā)現(xiàn)在病原細(xì)菌-野油菜黃單胞菌感染植物的過(guò)程中,VgrS的受體組氨酸激酶發(fā)揮著關(guān)鍵作用。本論文研究從遺傳學(xué)、酶學(xué)和生物物理學(xué)方面提出了充分的證據(jù),證明VgrS突變或失活后會(huì)導(dǎo)致細(xì)菌鐵代謝紊亂的作用,這對(duì)于細(xì)菌適應(yīng)脅迫環(huán)境的重要意義。同時(shí)研究結(jié)果也提示以病原細(xì)菌的重要受體作為分子作用藥靶時(shí),在特定生存環(huán)境中反而有可能提高細(xì)菌的適應(yīng)能力。研究過(guò)程中為了獲得VgrS的受體細(xì)菌的生長(zhǎng)特性的影響,應(yīng)用了丹麥Biosense微生物生長(zhǎng)動(dòng)態(tài)監(jiān)測(cè)系統(tǒng)測(cè)試了不同類型的細(xì)菌及細(xì)菌突變體在NYG培養(yǎng)基中的的生長(zhǎng)曲線圖。該Biosense微生物生長(zhǎng)動(dòng)態(tài)監(jiān)測(cè)系統(tǒng)能夠長(zhǎng)時(shí)間的同時(shí)平行測(cè)試多個(gè)類型的細(xì)菌菌株樣品的生長(zhǎng)曲線,并且所有的過(guò)程都是自動(dòng)化的,為研究人員提出關(guān)于組氨酸激酶VgrS蛋白對(duì)于細(xì)菌適應(yīng)脅迫環(huán)境的相關(guān)機(jī)理提供了重要的數(shù)據(jù)支持。這說(shuō)明全自動(dòng)生長(zhǎng)曲線分析儀能夠很好的了解不同類型的病菌的生長(zhǎng)情況,這能夠?yàn)檠芯咳藛T今后以病原細(xì)菌的重要受體作為分子作用藥靶時(shí),在特定生存環(huán)境中反而探索有可能提高細(xì)菌的適應(yīng)能力方面進(jìn)行更為全面、深入的研究。
相關(guān)新聞推薦
1、碳青霉烯類耐藥革蘭陰性菌聯(lián)合藥敏試驗(yàn)方法學(xué)、流程、優(yōu)缺點(diǎn)和結(jié)果判讀(三)
2、不同濃度巴西蘇木素對(duì)耐碳青霉烯類鮑曼不動(dòng)桿菌生長(zhǎng)及抑制作用(三)
3、不同涂布方法與平板放置時(shí)間對(duì)乳酸菌的平板菌落計(jì)數(shù)法的影響
5、生食大眼金槍魚(yú)中生物胺產(chǎn)生菌分離純化、菌落總數(shù)、生長(zhǎng)曲線及形狀鑒定(三)